3.1 Středoškolská biologie - úvod

Biologie na středoškolské úrovni vás měla seznámit se základní strukturou a stavbou živých organismů, jejich základním zařazením do skupin - systému rostlin a živočichů podle vzájemné podobnosti a evoluční příbuznosti. Měli byste znát a chápat základními principy jejich fungování a základní mechanismy, které jim umožňují chovat se jako živé - tj. přijímat a vydávat látky do okolí, přijímat a využívat energii, kterou potřebují na udržení uspořádanosti vlastního těla a jeho životní projevy jako je pohyb a dráždivost, v rámci kterých mohou reagovat na změny jejich vnějšího prostředí, schopnost rozmnožovat se a také - z dlouhodobého hlediska - schopnost vyvíjet se a měnit v rámci evoluce.
Součástí tohoto porozumění je i znalost vnitřního uspořádání rostlin a živočichů, vývoje a modifikací smyslových orgánů a také pochopení systému řízení základních procesů, které v živých organismech (také ve vyšších savcích a člověku) probíhají na biochemické úrovni, avšak ovlivňují a řídí i úrovně další: úroveň příjmu a výdeje energie živým organismem, úroveň reakcí organismu na změny prostředí, úroveň chování živého organismu a jeho reakcí vzhledem k okolí. Měli byste znát základy genetiky, dědičnosti a možnosti přenosu vlastností mezi jednotlivými generacemi organismů v závislosti na typu rozmnožování.

Pro základní pochopení fungování lesních ekosystémů musíte vědět jak fungují rostliny, jako nejdůležitější autotrofní organismy, na kterých je postaven základ potravních řetězců a tím i produkční možnosti ekosystému a jeho biodiverzita. Pro toto pochopení musíte mít představu o stavbě a fungování jednotlivých buněk, rostlinných pletiv, biochemického podkladu řízení procesů v rostlinách, což jsou základní znalosti, které vám připomene tato kapitola.
3.2 Buňka - stavba
Stavba buňky, jednotlivé organely
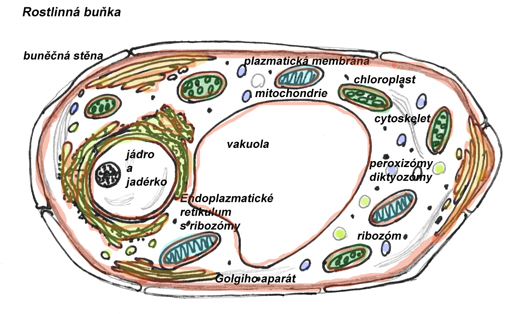
Rostlinná buňka je obklopená buněčnou stěnou na kterou na vnitřní straně navazuje biomembrána (plazmatická embrána, plazmalema). Uvnitř buňky je celá řada buněčných organel, které jsou rovněž obklopené biomembránou a mají v rámci buňky konkrétní funkci - oddělení biomembránou od zbytku cytoplazmy potom umožňuje průběh specifických chemických reakcí, udržení určitých podmínek vyšší nebo nižší koncentrace látek než ve zbytku cytoplazmy. Základním prostředím, které je podstatnou částí cytoplazmy i vnitřního prostředí všech organel a současně i prostředím buňky v rostlině obklopujícím, je voda.
Voda je optimálním prostředím pro biochemické reakce a to díky svým chemicko-fyzikálním vlastnostem. Voda je dobré polární rozpouštědlo - umožňuje příjem rozpuštěných minerálních látek, současně je rozpouštědlem i pro některé látky organické a díky malým molekulám, které ji tvoří má malou viskozitu a vysokou kapalnost a možnost prostupu membránami. Velká měrná tepelná kapacita - pomáhá udržovat stálou teplotu nejen jednotlivých buněk, ale i organismu jako celku. Vysoké měrné výparné teplo - napomáhá ochlazování organismů výparem vody- např. ochlazování listů při transpiraci (odpařením 1g vody dojkde k tepelné ztrátě 2450 J).
Buněčná stěna
Buněčná stěna je pevná struktura, která je typickou ochrannou povrchu a vnějším zpevněním buněk hub, rostlin a řas. Má funkci ochrannou a zpevňovací pro buňku, zpevňovací (a pomocí sřední lamely stmelovací) pro pletiva a tkáně mnohobuněčných organismů. Základní strukturní kostru buněčné stěny tvoří celulózy, hemicelulózy a pektiny. Kromě těchto základních látek existují další, kterými buňka zpevňuje buněčnou stěnu - některé bílkoviny, kutin, suberin, v případě hub chitin.
Základní struktura buněčné stěny je primární a sekundární buněčná stěna a na povrchu tzv. střední lamela jako rozhranní dvou sousedících buněk. Střední lamela je tvořená vzniká především z pektinových látek. Vnějším obalem buňky je primární stěna, vznikající jako první při vzniku nové a rostoucí buňky, obsahující zejména celulózy, hemicelulózy a pektiny, přičemž celulózová vlákna jsou v ní uspořádány v síťovině. Je pružná a snadno roste vkládáním nových celulózních mikrofibril, což je podstatné při prodlužování a růstu buněk. K primární stěně se po ukončení růstu buňky může zevnitř přikládat ještě sekundární stěna, která je zpravidla výrazně silnější, její celulózní vlákna jsou uspořádána rovnoběžně podél primární stěny - tloustnutí buněčné stěny, vedoucí k vyšší mechanické pevnosti buněk a pletiv (kolenchym, sklerenchym). V buněčné stěně se nacházejí malé otvory, kterými procházejí plazmodezmy - vlákna cytoplazmy spojující protoplasty sousedních buněk.
Biomembrána
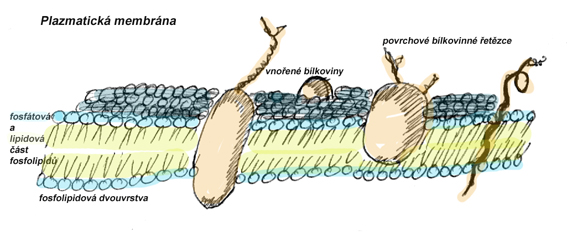
Biomemebrána je vnější polopropustný obal ohraničující buňku i její vnitřní organely. Má funkci ochrannou, transportní - umožňuje selektivní výdej a příjem látek, a informační (povrchové bílkovinné struktury identifikují např. vlastní a cizorodé buňky).
Skládá se z fosfolipidové dvouvrstvy a v ní ukotvených bílkovin. Fosfolipidová dvouvrstva, s proti sobě orientovanými hydrofobními lipidovými částmi molekul a směrem ven orientovanými hydrofilními částmi molekul, udává fyzikální vlastnosti plazmatické membrány: smáčivost, omezený prostup vody a látek rozpustných ve vodě, pružnost, plasticitu (používá se model tekuté mozaiky). Vnořené bílkoviny umožňují řízený transport látek; vytvářejí bílkovinné kanály, nebo přenašeče, se specificitou na vazbu určité látky.
Membránové organely
Endoplazmatické retikulum, Golgiho komplex (GA), vakuoly a ostatní endozomální struktury, vytváří obvykle vnitřní systém membrán, kterým je buňka dále členěna a umožňuje jí lepší organizaci složitějších životních pochodů.
Jednoduché membránové organely
Jsou to jednoduché měchýřky tvořené plazmatickou membránou, což vede k oddělení vnitřního prostředí měchýřku od cytoplazmy a dovoluje udržet v něm jiné prostředí než je v cytoplazmě (jiná acidita, koncentrace látek) a tím i oddělení chemických procesů v měchýřcích probíhajících. Může v nich docházet ke skladování meziproduktů metabolismu, likvidaci toxických látek, redoxním procesům... Podle typu chemických reakcí, které v nich probíhají se označují jako peroxizómy, glyoxizómy,
Endoplazmatické retikulum
Endoplazmatické retikulum (ER) je soustava vzájemně propojených miniaturních membránových systémů , tzv. cisteren a kanálků. Je v těsné blízkosti a napojení na buněčné jádro a obvykle i na Golgiho aparát. Diktyozómy jsou navzájem popropojované membránové útvary související s endoplazmatickým retikulem, které na straně přivrácené k jádru neustále přijímají váčky odštěpené od ER a intenzívně odštěpují váčky naplněné různými buněčnými sekrety. Rozlišujeme ER, na jehož vnějším povrchu jsou přisedlé ribozómy (drsné endoplazmatické retikulum), a ER bez ribozómů (hladké). Drsné endoplazmatické retikulum - s navázanými ribozomy - má význam především při syntéze strukturálních a enzymatických bílkovin. Ty jsou pak za pomoci ER a Golgiho komplexu transportovány do různých míst buňky. Hladké endoplazmatické retikulum propojuje hrubé ER a golgiho aparát a zajišťuje transport různých makromolekul mezi těmito dvěma systémy. Dále hraje významnou roli při syntéze lipidů, hormonů.
Golgiho aparát
Soustava Golgiho aparátu slouží k transportu, přechovávání a úpravě (tzn. posttranslační modifikaci) bílkovin (navazování a odstraňování nebílkovinných (zejména cukerných) složek, syntéza polysacharidů a imunoglobulinů a štěpení nebo transformace řetězců). V Golgiho aparátu se také tvoří polysacharidy, jako materiál pro tvorbu buněčné stěny.
Jádro a jadérko
Jádro je organela, v níž je uložena většina genetického materiálu (DNA) buňky. Má funkci nejen ochrannou pro uložení DNA, ale i řídící- pro většinu pocesů v buňce. Je organelou kde probíhá první fáze proteosyntézy (tvorby bílkovin). Prokaryota je název pro evolučně velmi staré organismy, které ještě nemají jádro oddělené biomembránou od cytoplazmy (patří tam např. bakterie). Všechny další, vývojově vyšší organismy už jádro oddělené biomembráno od cytoplazmy mají a nazývají se eukaryota (a patří tam rostliny, živočichové, houby, prvoci, …)
Jádra se často přizpůsobují i tvaru buňky. V typickém případě jsou zřejmě jádra buněk kulovitého či mírně protáhlého tvaru. Je to útvar oddělený od cytoplazmy dvojitou biomembránou s póry tvořenými speciálními bílkovinami, zvětšujícími její prostupnost pro molekuly RNA. Ta je na vnitřní straně vyztužena systémem intermediárních filament, tzv. jadernou laminou. Uvnitř jádra se nachází karyoplazma, tedy vlastně cytoplazma s charakteristickým jaderným složením. Uvnitř se nachází chromatin, tedy DNA a různé přidružené bílkoviny, ale i další struktury (např. jadérko), kde probíhají různé enzymatické procesy související s DNA a RNA a jejich tvorbou, replikací a postsyntetickými úpravami.
Jadérko je malá organela kulovitého tvaru uvnitř jádra, která obsahuje velké množství ribozomální RNA (rRNA). Vyskytuje se ve většině eukaryotických buněk. Jadérek bývá v typickém případě v jedné buňce jen několik, ale může dojít k jejich namnožení do výrazně vyšších počtů. Dále se v jádře zřejmě nachází rozvinutá síť cytoskeletu, která je tvořen zřejmě především mikrofilamenty a intermediárními filamenty.
Semiautonomní organely
Semiautonomní organely jsou buněčné organely s vlastní genetickou informací, a to v DNA v kruhovém uspořádán. U těchto organel se (vzhledem k dvojité biomembráně a tomuto typu DNA ) předpokládá, že vznikly na základě symbiózy mezi předkem eukaryotických organismů a organismy prokaryotickými. Semiautonomní – částečně autonomní jsou proto, že jsou schopné vlastní syntézy bílkovin a mají vlastní DNA, avšak na druhou stranu podléhají řízení z jádra buňky a nejsou schopné samostatného fungování mimo ni. Jsou od zbytku buňky odděleny dvojitou cytoplazmatickou membránou tvořenou dvojnásobnou lipidovou dvouvrstvou (předpokládá se, že vnitřní membrána z dvojice představuje původní cytoplasmatickou membránu původní bakterie nebo sinice a vnější membrána představuje původní cytoplasmatickou membránu hostitelské buňky). Obě membrány mají částečně odlišné vlastnosti, ať v již zastoupení membránových proteinů a zastoupení odlišných lipidů. Probíhají v nich procesy energetického metabolismu buňky. Patří sem mitochondrie, které mají svůj původ v bakteriích a jejichž funkcí je buněčné dýchání, a plastidy (chloroplasty), jejichž předchůdcem jsou zelené sinice, nebo řasy (případně výjimečně zelené řasy) a které mají hlavní funkci při fotosyntéze, případně při přechovávání barviv či zásobních látek fotosyntézou vzniklých. Dalším parametrem, kterým je možno popsat jakékoliv jádro, je počet chromozomových sad, které se v nich nalézají. U člověka jsou haploidní pouze pohlavní buňky, vajíčko a spermie, u rostlin všechny buňky gametofytu. Pro většinu buněk eukaryot je nicméně typické mít v jádře dvě sady genetické informace, tedy být diploidní. Z důvodu umožnění transportu mRNA k ribozómům a postranslační úpravy bílkovin je jádro napojeno na drsné endoplazmatické retikulum.
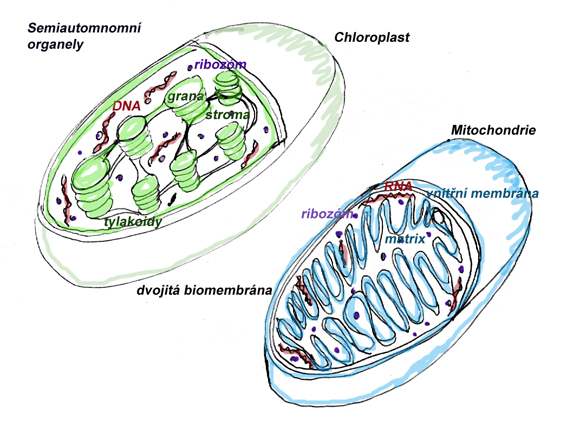
Mitochondrie
Vnitřní membrána je místem vlastní oxidace a je členěna v kristy a tvoří tak oddělené prostory (kompartmenty) pro různé chemické reakce a zároveň tím zvětšuje svou reakční plochu. Vnitřní prostor se nazývá matrix. V mitochondriích probíhá oxidativní fosforylace – velmi efektivní způsob získávání energie štěpením cukrů až na oxid uhličitý a vodu. V mitochondriích se utváří velká většina buněčného ATP a představuje tak základní zdroj energie eukaryotické buňky.
-
Plastidy - Chloroplasty
Stejně jako mitochondrie mají dvojitou biomembránu a vlastní DNA a ribozómy. Podobně jako mitochondrie mají složitým membránovým systémem rozčleněný vnitřní prostor na grana uspořádaná v tylakoidech a vnitřní stroma. Plastidy se vyskytují u několika skupin organismů, volně patřících k rostlinám: červené a zelené řasy, rostliny, sinice. U zelených řas se v buňce vyskytuje jen jediný plastid, u vyšších rostlin počet plastidů na buňku není vázaný, dělí se nezávisle na buněčném dělení a bývá jich v jediné buňce mnoho. Plastidy můžeme dále dělit podle obsahu barviv: Protoplastidy – Nejsou plně vyvinuté, nemají dostatečný obsah chlorofylu. Chloroplasty – Obsahují zelené barvivo chlorofyl. Mají plně vyvinutý fotosyntetický aparátem, představují probíhá v nich fotosyntéza. Chromoplasty – Obsahují jiný typ barviv než chlorofyl - karotenoidy, xantofyly.., mají obvykle ochranou funkci a také zabarvují rostlinu (např. podzimní listí). Leukoplasty – Neobsahují barviva, ale produkty fotosyntézy, především škrob.
Cytoskelet
Cytoskelet tvořený aktinovými mikrofilamenty (mikrovlákny) a mikrotubuly udržuje její tvar a vytváří možnosti pro pohyb cytoplazmy a cílený pohyb čehokoliv uvnitř buněk.
3.3 Procesy v buňce
Příjem látek do buňky
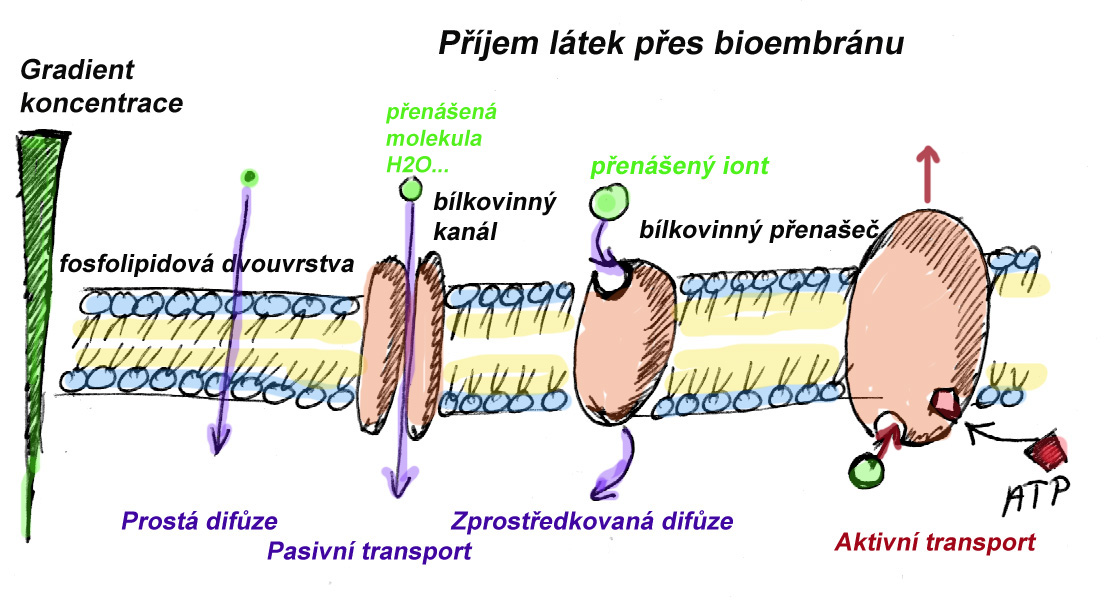
Pasivní
Přenos částice přes biomembránu ve směru koncenračního spádu - částice je přenášena z místa s vyšší koncentrací na místo s nižší koncentrací. Pro přenos není nutné dodat energii.
- Prostá difuze
Jedná se o nespecifický transport
Funguje na krátké vzdálenosti(v rámci buňky, mezi organelami a cytoplazmou, v rámci rostlinných pletiv). Rychlost záleží na rozdílu koncentrací přenášené látky na obou stranách biomembrány, na elektrickém náboji přenášené látky, velikost molekuly, propustnosti biomembrány dané teplotou apod. Probíhá buďto přímo přes fosfolipidovou dvouvrstvu (kyslík a některé další plyny a látky s velmi malými molekulami), nebo přes bílkovinné kanály (limitem je jen velikost molekul - např. transport vody). - Zprostředkovaná difuze
Funguje za pomoci bílkovinných přenašečů, které jsou specifické (schopné vázat jen určitý typ iontů). je rychlejší než prostá difůze, ale může zde fungovat kompetitivní inhibice mezi ionty různých prvků se stejným nábojem.
- Prostá difuze
Jedná se o nespecifický transport
-
Aktivní - zprostředkovaný transport
Přenos částice přes biomembránu proti koncentračnímu spádu, nebo tak, že její energie vzroste. Energie vzrůstá proto, že tam, kam se částice transportuje, má vyšší koncentraci, a/nebo tam má v důsledku membránového potenciálu vyšší elektrostatickou energii. Z tohoto důvodu musí tento typ trasportu být energeticky dotován - pomocí ATP, nejčastěji vazbou ATP na bílkovinný přenašeč, který tím změní strukturu, což mu umožní přijmout/nebo odevzdat přenášený iont. Může jít o ransport různě nabitých iontů současně- stejnými nebo opačným směrem. Celkový náboj přenášených částic z jedné a druhé strany membrány pak určuje, jestli se jedná o přenos elektroneutrální - nemění se elektrický náboj biomembrány (symport - současný transport dvou opačně nabitých iontů stejným směrem nebo současný transport stejně nabitých iontů opačným směrem - proti sobě), nebo transport elektrogenní, vedoucí ke změně elektrického potenciálu membrány.
- Primární aktivní transport,
např. sodíková pumpa - Na+/K+-pumpa, zajišťující protisměrný primární aktivní transport iontů Na+ a K+. Na úkor využití energie jedné molekuly ATP transportuje 3 ionty Na+ z buňky a 2 ionty K+ do buňky. Je běžná na všech biomembránách, protože nerovnováha intra- a extracelulárních koncentrací bazických iontů patří k základním znakům živých buněk - Sekundární aktivní transport
Na rozdíl od primárního aktivního transportu umožňuje i transport částic bez elektrického náboje (ovšem opět proti koncentračnímu spádu a s nutností dodání energie). Energie, potřebná pro tento transport, se získává z ATP a transportem elektricky nabité částice (např H+) se mění elektrický potenciál na mebráně, nebo gradient pH, což následně umožní souběžný transport částice bez elektrického náboje. Např. transport ATP a ADP (membrány tylakoidů, mitochondrií, …), transport glukosy spolu se sodnými ionty (symport)...
Vodíková pumpa – přenos H+ proti elektrochemickému gradientu, ATP odštěpí fosfát, změna pH a el. potenciálu, což může být využito pro transport dalších látek (sacharidy/H+, aminokyseliny/H+) Je běžná všech buňkách, ale ve větší míře zejména v kořenech. Umožňuje i regulaci pH, turgoru...
- Primární aktivní transport,
Metabolismus buňky
Základním jevem všech živých organismů je neustálá spotřeba energie pro udržení svých životních procesů a základního metabolismu. V buňkách probíhá neustále velké množství chemických reakcí, díky nimž dochází k tvorbě a přeměnám látek, tedy metabolismu. To umo6ňuje udržovat stálé vnitřní prostředí, opravovat a upravovat poškozené části organel, umožnit růst a řízení buňky a její reakce na vnější prostředí. Skladné procesy metabolismu, vedoucí k syntéze složitějších organických látek nebo složitějších buněčných struktur, se označují jako anabolické, rozkladné procesy, vedoucí k rozkladu organických látek a často za současného uvolnění energie, jsou tzv. katabolické. Obvykle jsou metabolické dráhy řízeny enzymaticky, tzn. pomocí látek, které katalyzují tyto reakce. Princip řízení je v zásadě z jádra buňky, protože katalyzující enzymy mají bílkovinnou část molekuly vytvářenou proteosyntézou.
Základním anabolickým procesem je fotosyntéza, probíhající u fotoautotrofních organizmů (sinice, rostliny a řasy). V světelné fázi fotosyntézy dochází za poutáním sluneční energie k tvorbě NADPH a ATP, v temnostní fázi jsou za pomoci těchto látek syntetizována v Calvinově cyklu z oxidu uhličitého a vody glukóza. Naopak základním katabolickým procesem je buněčné dýchání (respirace), při níž se rozkládají energeticky bohaté organické látky za vzniku ATP (a uvolňuje se oxid uhličitý).
3.4 Dělení buňky
K buněčnému dělení dochází v průběhu života – je to opravný mechanismus pro poškozené nebo odumírající buňky . V živých tkáních k němu dochází různě často podle rychlosti opotřebování dané tkáně. Detaily buněčného cyklu se liší podle druhu organismu, ale základní úkol dělení buněk je vždy stejný – zkopírovat svoji genetickou informaci a přenést ji na další generaci buněk. Nejjednodušší a nejrychlejší způsob buněčného dělení u prokaryotních organismů, které nemají ohraničené jádro a obsahují pouze jeden chromosom. Buněčné dělení u eukaryot je mnohem složitější, protože mají genetickou informaci rozdělenou do mnoha chromozómů a v rámci buňky je třeba replikovat i řadu buněčných organel. Cyklus buněčného dělení sestává nejen z fází, kdy se buňka skutečně přímo dělí, ale z časově mnohem náročnějších přípravných fází,kdy dochází ke shromažďování a syntéze látek, které budou při dělení třeba a hromadění energie ve formě zásobních látek.
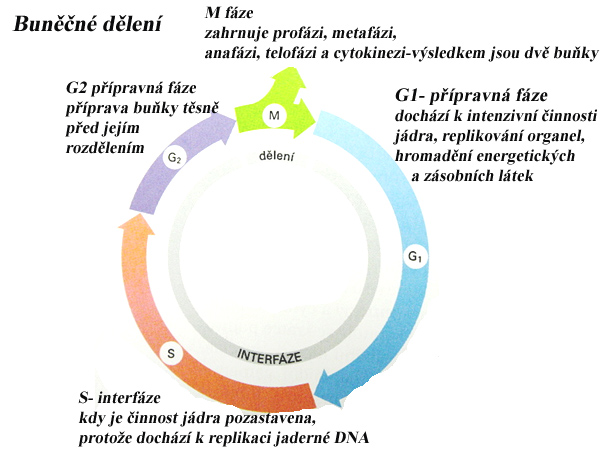
Mitóza
Z jedné diploidní mateřské buňky, vznikají dvě naprosto stejné buňky dceřiné, také diploidní. Dělí se na 5 fází – profáze, prometafáze, metafáze, anafáze a telofáze Před začátkem mitózy byl každý chromosom zreplikován a je tvořen dvěma identickými sesterskými chromatidami. Ty jsou spojeny proteiny na povrchu obou chromatid které se v průběhu mitózy štěpí, sesterské chromatidy se od sebe oddělují a vznikají dceřiné chromosomy, které jsou taženy mitotickým vřeténkem k opačným pólům buňky.
- Profáze: Ke konci S-fáze došlo ke zdvojení centrosomu, vně jádra začíná vznikat mitotické vřeténko interakcí mikrotubulů, dochází ke kondenzaci zreplikovaných chromosomů. Jaderný obal se rozpadá. Každá chromatida má na centromeru vázány proteinové komplexy (kinetochory). Mikrotubuly vřeténka se navazují na kinetochory.
- Metafáze: Chromosomy se uspořádají v ekvatoriální rovině (poloviční vzdálenost mezi oběma póly vřeténka), a vytvoří metafázovou destičku.
- Anafáze: Na začátku je spojení mezi chromatidami přerušeno enzymy, což umožňuje každé chromatidě (nyní dceřiné chromozomy) pohybovat se k pólu svého vřeténka. Což vede k rozdělení chromosomů do dvou identických sad.
- Telofáze: Kolem každé sady chromosomů se tvoří nový jaderný obal a vznikají dvě dceřiná jádra. V rámci cytokineze dochází k vytvoření přehrádky v buňce (cytoplazmě) a jejímu rozdělení). U živočišných buněk se vytváří dočasný kontraktilní prstenec, který se po skončení cytokineze opět rozpadá. U rostliných buněk vzniká uvnitř buňky nová buněčná stěna, obklopená membránou. Pozice stěny určuje polohu obou dceřiných buněk k okolí a spolu s růstem buňky i konečný tvar rostliny.
Meióza
Meióza (meiotické dělení, redukční dělení) je buněčné dělení, během kterého dochází k produkci buněk se zredukovaným (polovičním) počtem chromozómů, což je základní proces umožňující pohlavní rozmnožování.Haploidní pohlavní buňka (poloviční počet chromozómů) po splynutí s druhou haploidní buňkou vytvoří buňku diploidní, ze které pak mohou další meiozou vznikat somatické diploidní buňky. Meióza sestává ze dvou po sobě následujících dělení, kdy z jedné diploidní buňky rodičovské vznikají 4 haploidní buňky dceřinné. Během prvního (heterotypického) dělení dochází k redukci počtu chromozómových sad, dceřiné buňky mají jen polovinu chromozómů oproti rodičovské. Následně se pak dceřinné buňky dělí znovu, tentokráte (homeotypicky - mitóze podobně).
Profáze I.
první meiózy, je oproti profázi mitozy (a druhé meiozy v rámci dělení) mnohem komplikovanější a dělí se na následující stádia:
Leptotene, kdy dochází ke despiralizaci vláken DNA a oddělení a zviditelnění chromozómů.
Zygotene, kdy se odpovídající párové chromozomy přibližují k sobě a spojují v oblasti centromery.
Pachytene, kdy dochází k vytvoření tzv. tetrád, a kdy může dojít ke crossing-overu čili výměně odpovídajících částí sesterských vláken (umožnění genetické variability v populaci).
Diplotene, během které se uvolní bílkovinné vazby mezi homologickými chromozomy a dochází k jejich postupnému oddalování.
Diakineze vede k rozchodu homologických chromozomů.
Metafáze I
Probíhá téměř současně s diakinezí, dochází k napojení vláken dělícího vřeténka z opačných pólů buňky na centromery chromozomů. Chromozomy se před seskupují ve středové -ekvatoriální- rovině buňky.
Anafáze I
Dvouchromatidové chromozomy se rozcházejí k opačným pólům buňky - jsou přitahovány pomocí dělícího vřeténka.
Telofáze I
Vzniká jaderná membrána a jadérko, u protilehlých pólů buňky se seskupují haploidní sady chromozomů, dělící vřeténko mizí. Buňka se postupně zaškrcuje a dělí se na dvě dceřiné buňky - organely do buněk se dělí náhodně. Následuje zaškrcení a rozdělení buňky (cytokineze).
Homotypické dělení
Homotypické dělení (resp. Meióza II, Zrací dělení ekvační) je v podstatě shodné s mitózou, jen k němu dochází za haploidní sady chromozomů. Dělí se na profázi II, metafázi II, anafázi II a telofázi II. Její průběh je velice blízký mitóze.
3.5 Rostlina - anatomická stavba
Kořen
Kořenový systém je součást rostliny, která má několik funkcí: kotví rostlinu v pevném podkladu,
umžňuje jí příjem vody a látek ve vodě rozpuštěných, umožňuje jí určitým způsobem ovlivňovat příjem těchto látek zěmnou charakteru půdního prostředí v těsné blízkosti kořene (např. vodíková pumpa),
je to část rostliny, ve které se tvoří fytohormony řídící určité procesy v rostlině. Většinou je to orgán podzemní, neschopný fotosyntézy.
Pozměněné kořeny mohou být použity určitými druhy rostlin také jeko zásobní orgán, orgán umožňující dýchání (tisovce, mangrovy),
orgán umožňující vychytávání vzdušné vlhkosti (epifyty). Možná funkce kořene je i při symbióze
(kromě dalších druhů rostlin i s např. houbami nebo bakteriemi) a u některých druhů s vegetativním rozmnožování.
Jednotlivé části kořenového systému mohou být specializované:
kořeny kotevní, kořeny umožňující příjem vody, jemné kořeny v povrchové části půdy umožňující příjem živin.
Anatomická stavba kořene
Anatomická stavba kořene obsahuje různé druhy pletiv: na vnějším povrchu je rhizodermis – pokožka (jednovrstevné krycí pletivo), ze které vybíhá kořenové vlášení (jemné jednobuněčné útvary). Od ní směrem do středu kořene se nachází primární kůra ( exo-, mezo- a endodermis) a střední válec s cévními svazky, oddělený od primární kůry pericyklem. Cévní svazky ve stonku mohou mít různé typy uspořádání, v kořeni jsou většinou radiální. Druhotné tloustnutí kořene je možné u druhů, které mají mezi dřevní a lýkovou částí přístupné kambium.
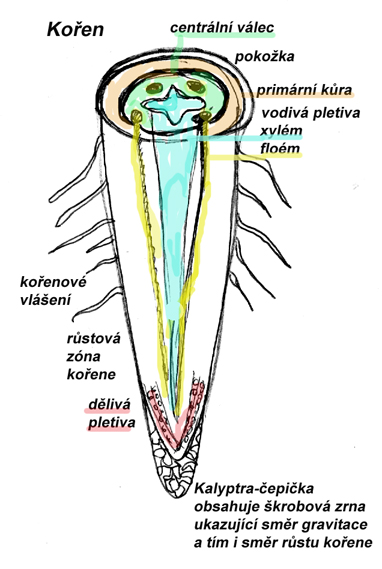
Morfologická stavba kořene
Vnější struktura kořene je určená jeho větvením a různými zónami s následujícími funkcemi:
kořenovou špičku kryje kořenová čepička, tzv. kalyptra,
což je pletivo kde tlak škrobových zrn na spodní část buněčné stěny umožňuje rostlině
(pomocí tvorby fytohormonu auxinu) růst geotropicky- tj. ve směru gravitace, do půdy.
Pod kalyptrou je růstová zóna kořene, s vrcholovými meristémy (dělivými pletivy) a řadou
nediferencovaných buněk. Nad touto zónou je zóna prodlužovacího růstu kořene a dále pak
část kořene diferenciační, s jemným kořenovým vlášením a maximálním příjmem živin.
Podle funkce je možné dělit kořeny na kotevní, upevňující rostlinu v půdě, a kořeny, které jí umožńují přijímat vodu, a povrchové kořeny podstatné pro příjem živin. Kořeny jsou uspořádány do tzv. kořenového systému, který je druhově specifický a může obsahovat různé proporce jemných a silných kořenů. Rozeznáváme kůlový kořenový systém (při jasné dominanci jednoho k kořenů), systém srdčitý (kotevní úlohu má více kořenů systému, kořenový systém povrchový…
Modifikace kořene
kořenová hlíza jako zásobní orgán, bulva, která má podobný vzhled jako kořenová hlíza, avšak část orgánu tvoří i spodní část stonku a tím pádem jsou zde i pupeny, příchytné kořeny sloužící k uchycení k pevnému odkladu (i mimo půdu), haustoria - sloužící k uchycení parazitické rostliny v tkáních hostitele a příjmu vody a živin, vzdušné kořeny umožňující příjem vzdušné vlhkosti, opěrné nebo chůdovité kořeny, sloužící ke stabilizaci dřevin v zamokřeném terénu, asimilační kořeny (u rostlin, které nerostou v půdě ale jako epifyty- např. orchideje).
Funkce kořene související se symbiózou se mohou vztahovat k symbióze s tzv. hlízkovitými bakteriemi
(symbiotické fixaci dusíku) což je časté jen u některých druhů rostlin nebo dřevin,
Nebo k symbióze s houbou, což je jev častější a nazývaný mykorrhiza.
Mykorrhiza představuje vztah houby a cévnaté rostliny a
podle současných poznatků mykorhizu tvoří většina rostlin, nemá ji jen asi 5 % druhů (vodní a mokřadní rostliny,
ppř. ruderální - Urticaceae, Polygonaceae, Brassicaceae).
Přínos rostliny: energetické zdroje a organické živiny (v první řadě sacharidy, resp. obecně sloučeniny uhlíku,
též některé vitamíny)
Přínos houby: přísun vody a minerálních látek (zejména nahrazuje-li kořenové vlásky), zásobování fosforem příp.
též dusíkem (obdobně, rozklad proteinů a aminokyselin) a draslíkem
Obecně mykorrhiza umožňuje vyšší adaptabilitu dřevin na extrémní podmínky, odolnost proti kyselému spadu a těžkým kovům v půdě.
Stonek
Stonek (podle stavby stélka, stéblo, lodyha) je část rostliny spojující kořenový systém s produkční a reprodukční částí
rostliny. Na stonku nacházíme listy, květy, pupeny. Funkcí stonku je spojovat fotosyntetický aparát (listy) s kořenovým systémem
a rozvádět v rámci rostliny vodu s rozpuštěnými minerálními látkami z kořenů a opačným směrem asimiláty z listů v rámci rostliny i do kořenů.
Zvyšování biomasy stonku umožňuje rostlině výškový růst a tím i větší konkurenceschopnost v kompetici o světlo.
Další vlastností stonku může být i schopnost fotosyntézy a druhotné tloustnutí (dřevnatění).

Anatomická stavba stonku
Stonek je kryt pokožkou - epidermis,která kryje primární kůru tvořenou parenchymatickým pletivem
a v centrální části stonku jsou vodivá pletiva- xylém a floém v různém uspořádání v cévních svazcích.
Cévní svazky bývají zpevněny sklerenchymatickými vlákny, zajišťujícími celkovou pevnost stonku.
Růstové zóny kořene jsou ve vrcholovém meristému a vmezeřených- interkalárních meristémech.
Krom toho dřevnatějící stonky mají charakteristicky sekundární meristém - kambium - umožňující diferenciaci nových vrstev xylému a floému a vytvářející u dřevin v našich podmínkách letokruhy.
Větvení stonku je v zásadě dvou typů - vidličnaté, kdy není možné rozlišit hlavní růstový vrchol (obě větve mají stejný význam) a postranní větvení - kdy vedlejší větve jsou kratší a dosahují menší výšky než hlavní vrchol (apikální dominance a tzv. monopodiální větvení), nebo naopak vedlejší větve hlavní vrchol výškově přerůstají (sympodiální větvení).
Morfologická stavba stonku
Stonek obsahuje podobně jako kořen zóny prodlužovacího růstu, zvaná internodia
a zóny s dělivými pletivy, ze kterých může docházet k diferenciaci listů, postranních výhonů, pupenů, květů- tzv. nódy.
- Stéblo - dutý stonek trav, který má výrazné zóny s vyrůstajícími listy (kolénka)
- Stvol - bezlistý stonek vyrůstající z přízemní listové růžice
- Lodyha - stonek nesoucí listy v různém uspořádání
- Kmen - druhotně tloustnocí stonek dřevin
Charakter stonku může být rozdílný podle různého charakteru růstu rostliny: vzpřímený, poléhavý (s možností zakořenit z nódů), plazivý, ovíjivý a popínavý(liánovitý charakter růstu podle opory).
Modifikace stonku
Dužnatý stonek (např jako zásobárna vody - sukulenty a kaktusy)
Stonková hlíza - zásobní orgán, ztlustlá nadzemní část stonku
Oddenková hlíza - zásobní orgán, časté zejména u podzemních částí stonku (hlízy brambor)
Oddenek - odzemní většinou oléhavý stonek, umožńuje rostlině vegetativní množení a šíření (pýr)
Úponek - k uchycení na pevné podpoře (vinná réva)
Trn - ochranná modifikace proti konzumentům
List
Listy jsou orgány rostlin přizpůsobené maximálnímu zachytávání světla při fotosyntéze. Jsou tedy orgánem, který obsahuje nejvíce fotosyntetizujících pletiv a současně i struktur, umožňujících fotosyntézu jako takovou - umožňujících příjem oxidu uhličitého (pomocí průduchů) a transport vody, minerálních živin a vznikajících asimilátů. Další funkcí listů je transpirace - tedy fyziologický výpar vody umožňující pohyb transpiračního proudu- tedy transport vody a minerálních látek v rostlině a současně sloužící k ochlazování (výparem).
Anatomická stavba listu
zachycuje různé druhy pletiv: na vnějším povrchu je kutikula, vosková vrstva kryjící epidermis – pokožku (jednovrstevné krycí pletivo bez chloroplastů). Součástí epidermis jsou průduchy a případně pozměněné buňky epidermis vytvářející strukturovaný povrch (např.trichomy). Pod pokožkou je vrstva palisádového parenchymu (protáhlé buňky s velkým počtem chloroplastů). U listů s výrazně odlišenou svrchní a spodní stranou je palisádový parenchym pod horní pokožkou a průduchy na spodní straně listu, u listů bez tohoto rozlišení je palisádový parenchym pod oběma pokožkami a průduchy rovněž součástí obou. Vnitřní stavbu listu tvoří houbový parenchym (velké buňky s výraznými mezibuněčnými prostorami umožňující transport vody a plynů mezi podprůduchovou dutinou, cévními svazky a palisádovým parenchymem), který obklopuje cévní svazky s vodivými pletivy.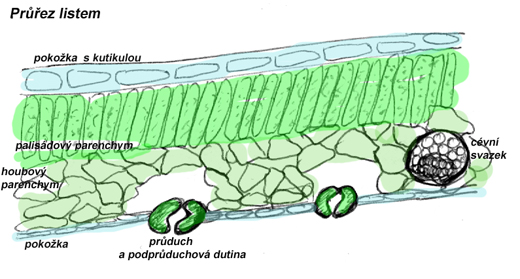
Průduchy
- U většiny rostlin jsou průduchy pouze na spodní straně listů, výjimku tvoří jehličnaté dřeviny, rostliny s listy postavenými kolmo - například trávy, a plovoucí vodní rostliny.
- počet většinou 30-350 na 1 mm2
- otevírají se po rozednění,
při snížené koncentraci CO2 v mezibuněčných prostorách,
reagují na modré záření(pravděpodobně receptory ve svěracích buňkách) - zavírají po západu slunce,
při nadměrné ztrátě vody,
při nízké vlhkosti vzduchu,
při vyšší koncentraci osmotika u kořenů (půdní sucho)se v kořenech tvoří ABA (kyselina abscisová), která podporuje uzavírání průduchů
Cévní svazky
jsou obklopené sklerenchymatickými (nebo parenchymatickými) pochvami a v listech tvoří žilnatinu (která je i charakteristickým rozlišovacím znakem). Ta je tvořená hlavními žilkami, spojenými menšími odbočkami (anastomózami). Podle charakteru větvení rozlišujeme:žilnatinu zpeřenou (charakteristická je jedna hlavní žilka, která prochází středem listu a z ní odbočují žilky postranní, které se dále větví),
znoženou (z báze čepele vybíhá jediná hlavní žilka, která se větví tak, že boční žilky vznikají pouze na jedné straně ),
dlanitou (z báze listu od řapíku vystupuje několik hlavních žilek),
souběžnou (souběžně vedených několik hlavních žilek od řapíku k vrcholu listu, typické pro jednoděložné),
rovnoběžnou (rovnoběžně vedených několik hlavních žilek které mají různou délku podle tvaru listu- např. u trav).
Morfologická stavba listu

Cévní svazky a žilnatina většinou souvisejí s tvarem listu a tvarem čepele. Podle základní stavby odlišujeme řapík a čepel listu. Řapík umožňuje přisednutí listu na stonek a obsahuje zejména cévní svazky. Listy bez řapíku jsou buď přisedlé nebo objímavé.
Čepel je buď jednoduchá nebo složená z několika lístků. Jednoduchá čepel
je buď celistvá nebo nejrůznějším způsobem členěná. Podle tvaru a členitosti čepele je celistvý list eliptičný,
srdčitý, vejčitý, obvejčitý, okrouhlý, kosočtverečný, kopinatý, trojúhelníkovitý, čárkovitý, jehlicovitý, střelovitý...
Podle okraje čepele může být celistvý list celokrajný, pilovitý, vroubkovaný, kracovitý, zubatý a vykrajovaný.
Výrazněji členěná listová čepel může být podle tvaru a hloubky zářezu buď dlanitě nebo zpeřeně členěná
(pak rozlišujeme listy na laločnaté, pokud jsou zářezy do jedné třetiny čepele a list označujeme
jako peřenolaločnatý a dlanitolaločný.
Pokud zářezy dosahují jedné poloviny čepele listy označujeme jako peřenoklané a dlanitoklané.
Pokud zářez zasahuje do dvou třetin čepele hovoříme o listech peřenodílných a dlanitodílných a když zářez dosahuje
až k hlavní žilce, hodnotíme listy jako peřenosečné a dlanitosečné.
Listy složené mají čepel rozdělenou na samostatné lístky a podle uspořádání na centrální ose jsou buď zpeřené nebo dlanitě zpeřené(sudo nebo lichozpeřené) . Členění může být poměrně složité a některé druhy (např. kapradiny) mají třikrát i vícekrát zpeřené listy. Dlanitě složené listy jsou tvořené z paprsčitě uspořádaných lístků na konci řapíku. Podle počtu lístků jsou troj, čtyř- až mnohočetné.
Listy mají na stonku nebo větvičce určité uspořádání (v jedné nebo několika rovinách),
které je rovněž rozlišovacím znakem pro určení druhů.
Postavení listů v jedné rovině může být vstřícné nebo střídavé (řapíky přisedají na větvičku proti sobě (javor),
nebo střídavě, nebo na pomyslné spirále okolo větvičky, postavení listů může být i v několika rovinách
(dvou a více- po obvodu celé větvičky) tzv. listy přeslenité.
Způsob složení čepelí a mladých listů v pupenech označujeme jako vernace (složená, svinutá, řasnatá…). Vzájemná poloha listových základů v pupenu se nazývá listová estivace. Pokud jsou mladé lístky uspořádány spirálovitě, je to estivace svinutá. Jestliže se překrývají, hovoříme o estivaci střechovité. Je potřeba si uvědomit, že listy se diferencují z dělivých pletiv v růstových vrcholcích pupene a samy o sobě dělivé pletivo nemají.
Modifikace listu
Možné modifikace listu:
Na klíčních rostlinách děložní lístky (zásobní funkce),
šupiny (chránící pupen), listeny v květenstvích,
cibule- zásobní orgány tvořené listy,
trny, listové úponky
Květ
Květ je orgán rostliny sloužící k pohlavnímu rozmnožování (což umožňuje genetickou variabilitu a možnost přizpůsobení). Květ tedy obsahuje reprodukční orgány a v průběhu vývoje z něj pak vznikají plod a semena. Současně je to část rostliny, která je druhově typická a podle které jsou rostliny zařazeny do systému.

Morfologická stavba květu
- Květní obaly - morfologicky nerozlišené:okvětí nebo morfologicky rozlišené: kalich a koruna - tvořené okvětními lístky
- Pestík - samičí rozmnožovací orgán, sestávající z blizny, čnělky a semeníku
- Tyčinky - samčí rozmnožovací orgány, sestávající z nitky a prašníku, produkujícího pyl
- Nektária - v případě květů opylovaných hmyzem
Z hlediska přítomnosti nebo nepřítomnosti obou typů pohlavních orgánů rozlišujeme květy na:
- Jednopohlavné - mají jen jeden typ pohlavních orgánů- jen tyčinky, nebo jen pestík
- Oboupohlavné - květ obsahuje oa typy pohlavních orgánů, kde ovšem jejich umístění a čas dozrávání může ovlivnit existenci možnosti samoopylení (samosprašnosti)
Z hlediska přítomnosti samčích i samičích květů na rostlině rozlišujeme potom rostliny na :
- Jednodomé - mají jednopohlavné květy ale obou pohlaví na jedné rostlině(kukuřice)
- Dvoudomé - mají květy jen jednopohlavné a jen jednoho typu na jedné rostlině (líska, rakytník)
Hlavním rozlišovacím znakem u květů je souměrnost a počet rovin souměrnosti květu.
Květy souměrné podle jedné roviny, která prochází středem a rozděluje květ na dvě zrcadlově symetrcké roviny jsou květy zygomorfní (např. fialka).
Květy souměrné podle dvou základních rovin, které dělí květ na 4 stejné části jsou květy bisymetrické (např. srdcovka).
Květy, které lze rozdělit více rovinami na více identických částí jsou květy pravidelné - aktinomorfní , kde počet rovin souvisí často s počtem okvětních lístků (např. hořec, pryskyřník).
Květy, kterými nelze proložit žádnou rovinu souměrnosti jsou květy asymetrické.
Květenství
Jsou uspořádáním souboru květů do určitého útvaru, vyrůstajícího ze stejného vřetena.
Podle jejich uspořádání dělíme květenství na jednoduchá a složená , přičemž složená květenství mají v odstatě tři úrovně uspořádání (jedotlivé květy v květenství a květenství uspořádaná v určitém souboru květenství.
Jednoduchá květenství se dále rozlišují na:
- Vrcholičnatá květenství Boční větve nesoucí květy přerůstají hlavní vřeteno. Květy vykvétají od středu k okraji, nebo odshora dolů. Typy květenství jsou označovány jako: vrcholík, vijan, vidlan, vějířek, srpek, lichopřeslen, šroubel, svazeček,klubko
- Hroznovitá květenství Hlavní vřeteno přerůstá boční větve nesoucí květy. Květy vykvétají od okraje ke středu a odzdola nahoru. Typy květenství jsou označovány jako: hrozen, lata, okolík,chocholík, hlávka, úbor, klas, klásek, palice, jehněda
3.6 Rostlinná pletiva (tkáně)
Rostlinná pletiva jsou tvořená větším počtem buněk společného původu, se stejnou komplexní funkcí v rostlině a často i obdobnou (charakteristickou) stavbou. Tyto rostlinné buňky jsou spojené pomocí střední lamely a navzájem se od sebe liší množstvím a velikostí mezibuněčných prostor, tvarem buněk, tvarem a silou buněčných stěn, a případně dalšími specializovanými funkcemi buněk, jako je např. jejich schopnost dělit se.
Základní rozdělení rostlinných pletiv je podle jejich funkce, stavby a místa výskytu v rostlině.
1. Podle typů buněk
- Jednoduchá - tvořená jedním typem buněk tvořících souvislé pletivo
- Složená - tvořená více typy buněk, které se ve svých funkcích navzájem podporují (např. vodivá pletiva - tvořena souborem cévních svazků umožňující transport vody a minerálních látek a podpůrných buněk udržujících funkci buněk vodivých).
2. Podle způsobů vzniku buněk
- Pravá-vznikají spojením dceřiných buněk v rámci buněčného dělení.
- Nepravá - vznikají druhotným nahloučením volných buněk (pseudoparenchym a plektenchym).
3. Podle tvaru buněk
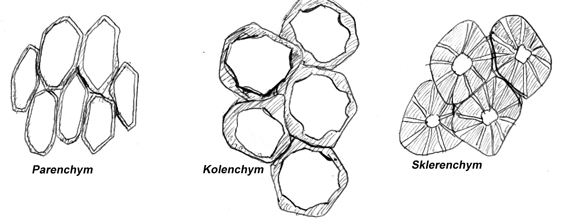
- Parenchym tenkostěnné buňky, mezi jejichž buněčnými stěnami jsou mezibuněčné prostory (interceluláry). Palisádový parenchym - jednosměrně mírně protáhlé buňky, kolmo k povrchu listu. S vysokým počtem chloroplastů v buňce je hlavním fotosyntetizujícím pletivem.Prozenchym se skládá z buněk jednosměrně protažených s šikmými přepážkami a bez mezibuněčných prostor(krycí pokožkové pletivo, vodivá pletiva). Aerenchym je silně rozvolněné řídké pletivo s oválnými buňkami, které se vyskytuje u vodních rostlin - hojné mezibuněčné prostory má vyplněny vzduchem. Hydrenchym je řídké pletivo s vysokou možností objemových změn sloužící např u kaktusů jako zásobárna vody.
- Kolenchym skládá se z tenkostěnných buněk se ztloustlou buněčnou stěnou v rohových spojeních nebo po stranách, což zvyšuje pevnost a současně i pružnost tohoto pletiva (je např. v řapíkách listů).
- Sklerenchym pevné pletivo, které má stěny buněk značně ztloustlé. Jednotlivé buňky se nazývají sklereidy a mohou být i bez živého obsahu. Má zpevňovací funkci(např.stěna pecky).
4. Podle funkce a schopnosti dělení na:
Dělivá - meristémy
- Primární
Protomeristémy a primární meristémy na růstovém vrcholku stonku a kořene a případně na vrcholových místech větvení stonku nebo v pupenech. Meristematické buňky se vyznačují větším jádrem, Endoplazmatickým retikulem a Golgiho aparátem. Buňky dělivých pletiv bývají menší a vakuoly většinou chybí. Vytvářejí buňky, které nejsou specializované a mohou se diferencovat na různé typy pletiv- což umožňuje růst a větvení, případně kvetení rostlin. Dále se dělí podle svého umístění na vrcholové, postranní a okrajové meristémy. - Sekundární
Vznikají obnovením dělivé funkce u trvalých pletiv a umožňují sekundární tloustnutí (zejména u dřevin). Nazývají se kambium a felogén a umožňují (z kambia) tvorbu dřeva - (sekundárního) xylému - směrem do středu a tvorbu lýka - (sekundárního) floému -od dělivého pletiva směrem k obvodu rostliny a z felogénu - obnovu borky-feodermu(zelenou kůru) směrem do sředu a k obvodu korek (felém).) . - Latentní
Jednotlivé buňky dělivého pletiva roztroušené na některých místech mezi buňkami trvalých pletiv. Aktivují se jen za určitých okolností a diferencijují adventivní kořeny a adventivní pupeny..
Trvalá
- Krycí Kryje povrch rostlinného těla a chrání rostlinu před škodlivými vlivy (epidermis, rhizodermis).
- Provzdušňovací Umožňují spojení rostlinných pletiv s okolím – především v rámci příjmu a výdeje oxidu uhličitého, vodních par. Patří tam průduchy (stomata – dvě svěrací buňky), čočinky (lenticely).
- Vylučovací Slouží k vylučování nebo hromadění různých produktů látkového metabolismu rostlin (voda, silice, pryskyřice, těkavé organické látky)- hydatoty, mléčnice, idioblasty, nektárie (medníky).
- Asimilační Slouží především k fotosyntéze a u rostlin mírného pásu je to především palisádový parenchym na svrchní straně listu pod pokožkou.
- Zásobní Mají velké vakuoly vyplněné zásobními látkami (škroby, tuky), běžně jsou v zásobních hlízách, zásobních orgánech
- Nasávací Kořenové vlášení, haustoria.
- Vodivá (viz níže).
Vodivá pletiva
Vodivá pletiva vznikají činností prokambia, látky se jimi přemisťují difúzí pomocí plazmodezem. U nižších rostlin nejsou vyvinuta, s prvními náznaky se setkáváme u mechorostů, u kapraďorostů jsou již plně vyvinuta.
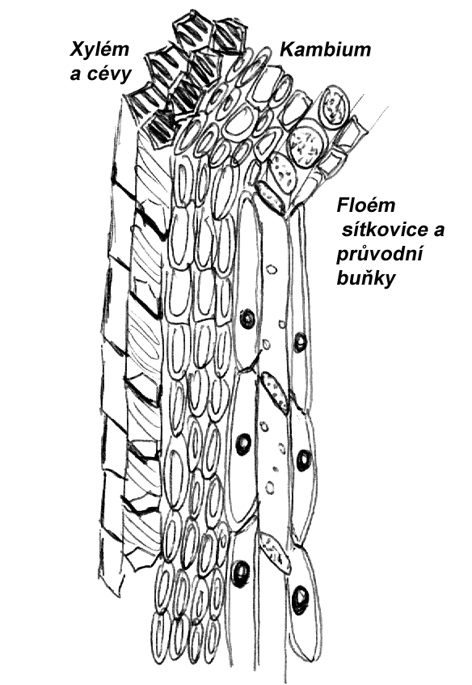
Prvotní dřevo - xylém (z řec. xýlon = dřevo)
Rozvádí vodu a roztoky anorganických látek od kořenů vzhůru. Tvoří ho čtyři typy buněk:- Cévice (tracheidy)
Úzké, trubicovité, na sebe navazující buňky, pěti- nebo šestihranné, jejichž příčné stěny nejsou proděravělé, a jež se během růstu stále více zešikmují. Mají nepravidelně ztlustlou buněčnou stěnu. V období činnosti jsou již mrtvé. - Cévy (tracheje)
Vznikly rozpuštěním přehrádek několika nad sebou stojících buněk bez živého obsahu (jsou tedy rovněž mrtvé), a vytváří široké trubice z protáhlých buněk s částečně nebo úplně rozpuštěnými přehrádkami. Scházejí nahosemenným a kapraďorostům, v buněčné stěně mají ztenčená místa, tzv. tečky. - Dřevní vlákna (libriform)
Tvořena sklerenchymem (z převážně mrtvých buněk) s mechanickou a zásobní funkcí. - Dřevní parenchym
Neprotáhlé, živé buňky, rozvádí vodné roztoky mezi obvodem a středem stonku (ve vodorovném směru).
Prvotní lýko - floem (z řec. phloiós = lýko)
Rozvádí produkty fotosyntézy z listů dolů, stejně jako dřevo ho tvoří čtyři typy buněk:
- Sítkovice
Vznikly spojením živých, do řad nad sebou uspořádaných protáhlých buněk s příčnými proděravělými stěnami označovanými jako sítka. Koncem vegetační doby se sítka ucpávají kalózou a sítkovice obvykle odumírají, některé rostliny na jaře ucpávky rozpouštějí (lípa). - Průvodní buňka
Protáhlá, těsně přiléhá k buňce sítkové, její jádro zůstává zachováno po celou dobu života buňky a ovlivňuje pochody v buňce sítkové, která své jádro brzy ztrácí. Někdy je buněk víc, u jehličnanů chybí. - Lýková vlákna a lýkový parenchym
Mají podobnou stavbu a funkce jako u dřevních vláken a parenchymu. Zajišťují mechanické zpevnění, parenchym rozvádí látky ve vodorovném směru. Buňky lýkového parenchymu mohou být dlouhé až 120 mm (např. len - využívá se toho v textilním průmyslu). - Sklerenchymatická lýková vlákna
Cévní svazky
Skládají se z části dřevní a lýkové. Dřevní část tvoří u kapraďorostů a nahosemenných cévice u krytosemenných cévice a cévy. Nejdůležitější částí lýka jsou sítkovice. Cévy i sítkovice jsou doprovázeny dřevním čil lýkovým parenchymem, prozenchymatickými a většinou i sklerenchymatickými buňkami, tzv. pletivy doprovodnými, používá se též název buňky průvodní. Od okolního pletiva jsou odděleny pochvou z parenchymatických buněk (osladič), která může zkorkovatět, nebo buňkami sklerenchymatickými (kukuřice).
V prýtu jednoděložných rostlin jsou cévní svazky roztroušeny, u dvouděložných jsou uspořádány do kruhu.
Cévní svazky uzavřené: vznikají přeměnou celého prokambia, druhotně netloustnou
Cévní svazky otevřené: zůstává zachována část prokambia, které se mění na kambium, svazky druhotně tloustnou
Typy cévních svazků:
- bočný (kolaterální) v lodyhách většiny rostlin
- dvoubočný (bikolaterální): po stranách dřevní část, uprostřed lýko, tykvovité a lilkovité
- paprsčitý (radiální): v kořenech
- soustředný
- lýkostředný: kapradiny
- dřevostředný: oddenky jednoděložných rostlin
3.7 Procesy v rostlině
Příjem vody rostlinou:
- -pasívní - difůzí
-aktivní - díky transpiraci - -přes kořenové vlášení do centrálního válce kořene - díky osmotickému tlaku
-xylémovou (dřevní) částí cévních paprsků - díky adhezním silám ( kapilaritě) a kohezním silám (přitažlivé síly mezi molekulami kapaliny) podporovaným transpirací
Hnací síly toku vody v rostlině:
1. Na krátké vzdálenosti:
-
Difůze
- hybnou silou je gradient koncentrace (pohyb molekul směrem k jejich nižší koncentraci. Při osmóze dochází k vyrovnávání koncentrací dvou roztoků o nestejné koncentraci přes polopropustnou membránu. Prostupují pouze molekuly vody směrem do místa s vyšší koncentrací rozpuštěných látek.
Typické jevy:
Plazmolýza: Stav buňky v hypertonickém roztoku, voda difunduje ven a dochází k odtržení plazmalémy od buněčné stěny
Plazmoptýza: Stav buňky v hypotonickém roztoku, voda difunduje do buňky a při příliš vysokém tlaku na vnitřní stěnu (turgor) dochází k jejímu poškození.
2. Na dlouhé vzdálenosti:
- Gravitační tok - ve floému, tok asimilátů podle gradientu teploty
- Transpirační proud - pohyb nepřerušeného vodního sloupce nasávaný podtlakem vzniklým výparem vody z listů podle gradientu teploty
- Podle gradientu hydrostatického tlaku - pod meniskem v kapiláře dochází ke snížení tlaku proti volné vodní hladině a tím k nasávání vody do kapiláry (průměr cévy 50 - 100 mikrometrů, mikrokapiláry až 10 - 4 mikrometrů)
- Kapilární elevace - jev vznikající díky povrchovému napětí vody,
umožňuje vodnímu sloupci vystoupit až do výšky 150 m (Sequoia, Pseudotsuga)
Objem vody proudící xylémem je mnohem větší než objem vody proudící floémem s asimiláty ke kořenům.
Transpirace
Hlavním určovacím procesem pro příjem vody je transpirace, což je výdej vody přes průduchy listů. K výparu dochází již uvnitř listů a výdej páry je regulován otevíráním průduchů (Zahrnuje více než 80 %, přes pokožku a kutikulu se odpaří jen 5 - 20 % vody). List může odpařit za 20 až 60 minut tolik vody, kolik sám váží. Výpar z listů je téměř stejný jako z volné vodní hladiny (15 metrů vysoký javor odpaří za hodinu 220 litrů). Jen 1 - 5 % vody, která rostlinou proteče, je použita k metabolismu. Význam transpirace spočívá především v ochlazování povrchu, dále má fyzikálně velký význam pro příjem rozpuštěných živin spolu s vodou přes kořeny.
Transpirační koeficient
počet molů vody vytranspirovaných při příjmu 1 molu oxidu uhličitého (u C3 rostlin 300 - 500, u C4 od 250)
Faktory ovlivňující transpiraci:
- Vnější - teplota, pohyb okolního vzduchu, vzdušná vlhkost, tvar povrchu, ze kterého se voda odpařuje
Teplota listu - pokud teplota vzduchu je nižší než 30°C, je osvětlený list teplejší. Je li teplota okolí vyšší než 30°C, je list chladnější, stejně jako v noci. Teplota malých listů se obvykle neliší od teploty okolí o více než 3 stupně. S rostoucí teplotou vzduchu musí růst i transpirace. (Osvětlený list v našich podmínkách absorbující 1/2 dopadající energie (1000W /m2) by se ohříval rychlostí 25 stupňů/min.) Na odpaření jednoho gramu vody je třeba 2450 J.
Tvar povrchu - na osluněném stanovišti je plocha listů obvykle menší, listy jsou více členěné
Vzdušná vlhkost - v prostředí s vyšší vzdušnou vlhkostí mají rostliny obvykle větší listy - vnitřní - pozice listů, stáří listu, počet a velikost průduchů, síla kutikuly, tlak vodní páry v mezibuněčných prostorech, tloušťka listu
3.8 Fotosyntéza

Je základní biologický proces zabezpečující život na zemi. Je to metoda získávání energie, kterou
využívají fototrofní organismy, především vyšší rostliny.
Z energie slunečního záření dopadajícího na Zemi je 40% odraženo a jen 1-5%
jsou využity rostlinami k syntéze glukosy z energeticky chudého oxidu
uhličitého a vody. Tvorba energeticky bohaté molekuly glukosy je navíc
provázena produkcí kyslíku, nezbytného pro život většiny organismů.
Fotosyntéza je v podstatě procesem přeměny energie fotonů v energii chemických vazeb v organických látkách.
Jde o dvojstupňový endogenní redukční proces, ve kterém je CO2 redukován vodíkem
pocházejícím z molekul vody za tvorby molekul glukosy.
Energie takto uložená
je pak v dalších enzymatických reakcích použita na tvorbu produktů fotosyntézy
- asimilátů, což jsou např. zásobní látky (škrob, bílkoviny, tuky a
jiné organické látky).
V podstatě všechny organické látky rostlinného těla
jsou tvořeny z vody, jednoduchých anorganických sloučenin přijatých kořeny a
oxidu uhličitého. 1 g asimilovaného CO2 odpovídá zhruba 0,4g sušiny rostlinného těla.
Proces fotosyntezy probíhá ve specializovaných organelách: chloroplastech
Chloroplasty: obsahují systém vnitřních membrán nazývaných thylakoidy.
Tyto uzavřené membrány stupňovitě na sebe naskládané tvoří grana
obkopená základní hmotou zvanou stroma. V granech jsou umístěny oba fotosystémy
I. a II. Chlorofyl je rozmístěn v membránách gran, mezi vnější bílkovinovou
a vnitřní lipidovou vrstvou.
1. Světelná fáze fotosyntézy:
Probíhá v granech chloroplastů, ve fotosyntetickém aparátu složeném z pigmentů (zejména chlorofylů),
které absorbují sluneční energii fotonů a uvolňují energeticky bohaté elektrony,
použitelné v dalších chemických přeměnách.
Rostliny mohou využívat spektrum v rozsahu 400 - 750 nm:
- chlorofyly a a b zachycují modrofialové a červené části spektra,
- fykocyanin a fykoerytrin (barviva vyskytující se u řas) zelené a žluté světlo,
- xantofyly a karotenoidy modrozelenou část spektra
Fotosystémy I a II jsou soubory anténního světlosběrného komplexhu a látek, přenášejících excitovaný lektron a tím i energii. Světlosběrné komplexy obsahují jako hlavní složky stovky (asi 300) molekuly chlorofylu a a b , které usměrňují energii fotonů k reakčnímu centru fotosysémů - molekulám a1 a a2. Ty se pak excitují, a díky vyšší energetické hladině odevzdávají elektron na celý řetězec jeho přenašečů. Záleží na konkrétním typu fotosystému (I. nebo II.) co je výsledkem a produktem tohoto přenosu.
Fotosystém I
Má absorpční maximum při 750 nm, a elektrony z reakčního centra jsou předávány přes systém přenašečů, přičemž výsledkem je tvorba NADP+ na NADPH a tvorba ATP - cyklická fosforylace (cyklická proto, že původně excitovaný elektron se po vrací zpět do centra fotosystému. ecyklická fosforylace vede k redukci NADPH a "únik" elektronu z ze systému. tyto chybějící elektrony jsou pak nahrazovány přísunem elektronů z fotosystému II.
Fotosystém II
Má absorpční maximum při 680 nm a elektrony z reakčního centra jsou využívány k fotolýze vody. Z ní je využit proton k redukci NADPH a elektron přenášen do reakčního centra fotosystému I. Vedlejším produktem je kyslík.
2. Temnostní fáze fotosyntézy - Calvinův cyklus (C3):
Probíhá paralelně s fází světelnou (není vázána na tmu!) ve stromatu chloroplastů.
a využívá molekuly ATP a NADPH, vzniklé ve světelné fázi k zabudování uhlíku do molekuly glukosy.
Prvotním akceptorem CO2 přijatého ze vzduchu je ribulosa 1,5 bifosfátu.
Proces se nazývá Calvinuv cyklus a jde o syntézu glukózy z oxidu uhličitého a vody, která probíhá zejména pod pokožkou listů, v palisádovém parenchymu a stromatu chloroplastů.
Energie k produkci glukózy je dodávána ve formě ATP a NADPH ,
energetických sloučenin vzniklých po absorpci fotonů ve světelné fázi fotosyntézy.
Ke tvorbě glukozy dochází zabudováním CO2 (přijatého průduchy ze vzduchu) do primárního
řetězce ve vazbě na primární akceptor ribulosa 1,5 bifosfát.
Calvinův cyklus využívají tzv. C3 rostliny, rostliny mírného pásu. Intenzita fotosyntézy u nich není příliš vysoká, ale nejsou tak náročné na intenzitu světla, snášejí i světelné výkyvy, nižší teploty a nižší dostupnost vody.
C4-cyklus: Hatch - Slackův cyklus (C4)
je další možností zabudování CO2 do uhlíkatého řetězce.
Používají ho především tropické a subtropické rostliny a fotosyntéza probíhá nejen v palisádovém parenchymu, ale i v buňkách obklopujících cévní svazky.
Rozdíl je v tom, že prvotním aceptorem CO2 je fosfoenolpyruvát
a prvotními produkty je malát, aspartát a oxalacetát.
Uhlík z těchto meziproduktů je pak přenesen na ribulosa 1,5 bifosfát
a přes 3- fosfoglycerovou kyselinu probíhají reakce dále stejně jako u Calvinova cyklu.
Fosfoenolpyruvát má několikanásobně větší afinitu k CO2 než
ribulosa 1,5 bifosfát.Což umožňuje fotosyntézu i při nižším množství
CO2 ve vzduchu a při omezeném příjmu vody, ale je potřeba vyšší intenzita slunečního záření.
Další výhodou tohoto způsobu je větší efektivita fixace a menší
ztráty při fotorespiraci a tím intenzivnější tvorba rostlinné hmoty.
Tento cyklus využívají takzvané C4 rostliny, rostliny tropů a subtropů. Mají poměrě vysokou fotosyntetickou kapacity, ale na snížení intenzity záření reagují jejím poklesem. Potřebují vyšší teplotu pro optimální průběh fotosyntézy.
Rozdíly mezi C3 a C4 rostlinami:| Charakteristika | C3 rostliny | C4 rostliny |
|---|---|---|
| Anatomie listu | houbový a palisádový parenchym | větší a silnější list s mohutnějším mezofylem, parenchym věnčitě kolem cévních svazků |
| Primární produkt vazby | C3 kyselina- fosfoglycerát | C4 kyselina - malát, oxalacetát, aspartát |
| Fotosyntéza mg CO2/dm2 plochy listu/hod | 15-40 | 60-100 |
| Teplotní optimum | 15-25°C min -6 až -10°C max 30°C |
30-40°C min pod +10°C |
| Potřeba pro fixaci 1 CO2 | 3ATP + 2 NADPH | 5ATP + 2 NADPH |
| Fotorespirace | až 1/3 fotosyntézy | velmi malá |
| Maximální rychlost růstu | 0,5 - 2g suš./1 dm2/den | 4 - 5g suš./1 dm2/den |
| Transpirace | 450 - 900g H2O/1 g sušiny | 250 - 350g H2O/1 g sušiny |
| Příklady | rostliny a dřeviny mírného pásu | kukuřice,lilkovité, cukrová třtina, bambus, merlíkovité |
CAM cyklus: cyklus tučnolistých (crasulacean acid metabolism)
Opět jde o modifikaci Calvinova (nebo přesněji Hatch-Slackova) cyklu. Zásadní modifikací je oddělení doby vázání oxidu uhličitého od přímého průběhu fotosyntézy. To má svůj význam pro rostliny z podmínek s vysokými denními teplotami a současně nedostatkem vody. To, že mohou mít přes den (za slunečního svitu, kdy probíhá fotosyntéza) zavřené průduhy, výrazně zlepšuje hospodaření s vodou a omezení výparu. Tyto rostliny (sukulenty a kaktusy) přes noc váží oxid uhličitý do malátu, který ukládají ve vakuolách (kyselý obsah vakuol, proto acid metabolism). Ve dne světelná část fotosyntézy probíhá zcela normálně, temnostní má jako substrát nikoli oxid uhličitý z atmosféry, ale oxid uhličitý vázaný na malát a přes něj transportovaný do Calvinova cyklu.
Faktory ovlivňující fotosyntézu:
Regulace fotosyntézy: Pomocí koncentrace asimilátů v buňceVnější:
- Světlo
- v červeném světle (660 nm, modré 460 nm) je intenzita fotosyntézy největší. Energie světelného kvanta je nepřímo úměrná vlnové délce, fotosyntéza má vyšší účinnost v červené části spektra než v modré. - Délka osvětlení
- Teplota
- optimum pro rostliny mírného pásu je okolo 20-25°C, nad 30°C fotosyntéza rychle klesá. - Obsah CO2 ve vzduchu
- příliš vysoký nebo příliš nízký fotosyntézu snižuje a může ji i zastavit (normální obsah O2 je 21%, CO2 0,035%) - Dostatek vody
- Přísun makro a mikroprvků:
Mg a Mn působí jako katalyzátory biochemických reakcí, S je součástí acetylkoenzymu A z Krebsova cyklu, P je používán pro fosforylaci, oxidoreduktasy fotosystémů obsahují Cu, Zn, Fe, N - součást bílkovin, K - regulace turgoru svěracích buňek atd.
- Světlo
-
Vnitřní:
- Druh, odrůda nebo ekotyp rostliny
- Množství a velikost chloroplastů v buňkách
- objem i povrch chloroplastů ve stínu je menší - Množství a velikost průduchů
- Postavení listů na stonku
(osvit), tvar listů - menší listy a větší rychlost proudění vzduchu zrychlují přenos vodní páry a CO2, tloušťka listů - ve stínu je menší - Stáří pletiv
- Fyziologický stav rostliny
Organické látky mohou být unášeny transpiračním proudem. Stromy hromadí v některých buňkách dřevní části na podzim škrob a na jaře je část cukru vzniklého z této zásobní látky přenášena k rašícím pupenům. Roztok, který je transportován floémem obsahuje obvykle 10-25% cukru. V xylému je ho stopové množství, ale na jaře koncentrace stoupá až na 2-3%. Protože objem xylému mnohonásobně převyšuje objem floému je množství šťávy transportované na jaře xylémem značné- a v některých případech i celkový objem cukru (př. cukrový javor). Když však listy vyraší a dospějí obsahuje šťáva v xylému jen stopové množství cukru.
Látky vzniklá fotosyntézou a obsažené v listech jsou před opadem odváděny do větví a kmene, ale zde jsou spotřebovány jen relativně málo, protože v té době jsou životní projevy již omezené. Přemisťování a ukládání asimilátů se může uskutečňovat jen v živých buňkách, především v parenchymu. Ve zdřevnatělém stonku je to především kůra, dřeňové paprsky a dřevní parenchym. Zásobní látky se zpravidla ukládají ve formě škrobu a na podzim ho bývá ve všech parenchymatických částech větviček maximální množství (možno vidět na mikroskopickém řezu po potření jódem). Ve starších větvích může být škrob v dřeňových paprscích a během zimy se může měnit na jiné formy zásobních látek. U lípy se ztrácí úplně, protože se mění na tuky a částečně na cukr. ale na jaře se opět objeví v kůře, dřeňových paprscích a dřevním parenchymu (různé u růz. dřevin). Na jaře jsou rašící pupeny živy ze zásobních látek, které mizí nejprve z rašících větévek, později i ze starších. Když jsou listy schopné krýt svou energetickou potřebu fotosyntézou je přísun živin do nich zastaven. To se děje postupně, s postupným dospíváním jednotlivých listů. Zásobní látky se ve výše uvedených pletivech začínají hromadit v okamžiku, kdy výkon fotosyntézy přesáhne energetickou potřebu listu a dále jsou výrazněji doplněny před opadem listů na podzim.
3.9 Řízení procesů v rostlině
Podstata vnitřního řízení procesů v rostlině, tak, aby fungovala nikoli jako soubor na sobě nezávislých buněk a pletiv, ale jako celistvý organismus, schopný reagovat na změnu podmínek vnějšího prostředí, je založený na přenosu chemických signálů, látek zvaných fytohormony. Jedná se o organické sloučeniny, syntetizované za určitých podmínek v různých částech rostliny. Dvě hlavní skupiny se, podle výsledků jejich působení v rostlině, označují jako stimulátory a inhibitory růstu růszných částí rostliny.
Stimulátory:
- Auxiny Stimulují tvorbu postranních - adventivních - kořenů a jejich prodlužování, stimulují apikální dominanci (růstovou převahu nejvyššího výhonku rostliny) a růst stonku, oddalují stárnutí a opad listů, květů a plodů. Zvyšují i dlouživý růst buněk (v souvislosti s gravitací se objevují ve spodní části jinak než vertikálně rostoucích větví).
- Cytokininy Podporují dělení buněk a tím i aktivitu meristémů. Stimulují růst kořenů, zakládání, tvorbu a růst postranních větví (odnoží, výhonků). Zpomalují stárnutí pletiv.
- Gibereliny Stimulují růst, zejména dlouživý růstu stonku. Hrají úlohu při klíčení semen, rašení a tvorbu pupenů a kvetení (jejich aplikací lze přerušit dormanci). Aplikací giberelinů lze přerušit dormaci semen i pupenů.
- Brassinosteroidy Mají význam zejména při kvetení, podporují růst pylové láčky (mají vysoký obsah v pylu), regulují kvetení a reprodukci, rovněž jsou obsaženy v semenech a ovlivňují jejich klíčení.
Inhibitory:
- Kyselina abscisová Patří mezi inhibitory růstu podporující nástup do a udržení dormance, opad listů, ukončení vegetační doby. Tvoří se v pupenech, mladých listech, semenech, kořenech. Je využívána i jako signální látka pro uzavření průduchů v době půdního sucha.
- Ethylen Zrychluje zrání plodů, opad listů, omezuje dlouživý růst. Vytváří se mimo jiné i v plodech.
Literatura použitá a doporučená:
- Gloser J., 1995: Fyziologie rostlin, PrF MU, Brno
- Novák J., Skalický M., 2008: Biologie: cytologie, histologie, organologie a systematika Praha, Powerprint
- Procházka S. a kol., 1998: Fyziologie rostlin. Praha : 484 s.
- Rosypal S., 1998: Přehled středoškolské biologie, nakladatelství Scientia, 534 s.
- Šebánek J. a kol.,1983: Fyziologie rostlin. SZN, Praha: 558 s.
- Volf F., 1988: Zemědělská botanika. SZN, Praha: 352 s.